Poster title: Functional specialisation of dendritic cell subsets and how dendritic cells initiate and shape adaptive immune responses
Author: Xin Xiong
Affiliation: College of Life Sciences, Sichuan University
Contact: Lucimorl123@gmail.com
Course context: BI Advanced Immunology poster and final presentation
This page provides supplementary information for the QR code on the poster, including figure captions, image/source notes, acknowledgements, selected references and a concise mechanistic research note.
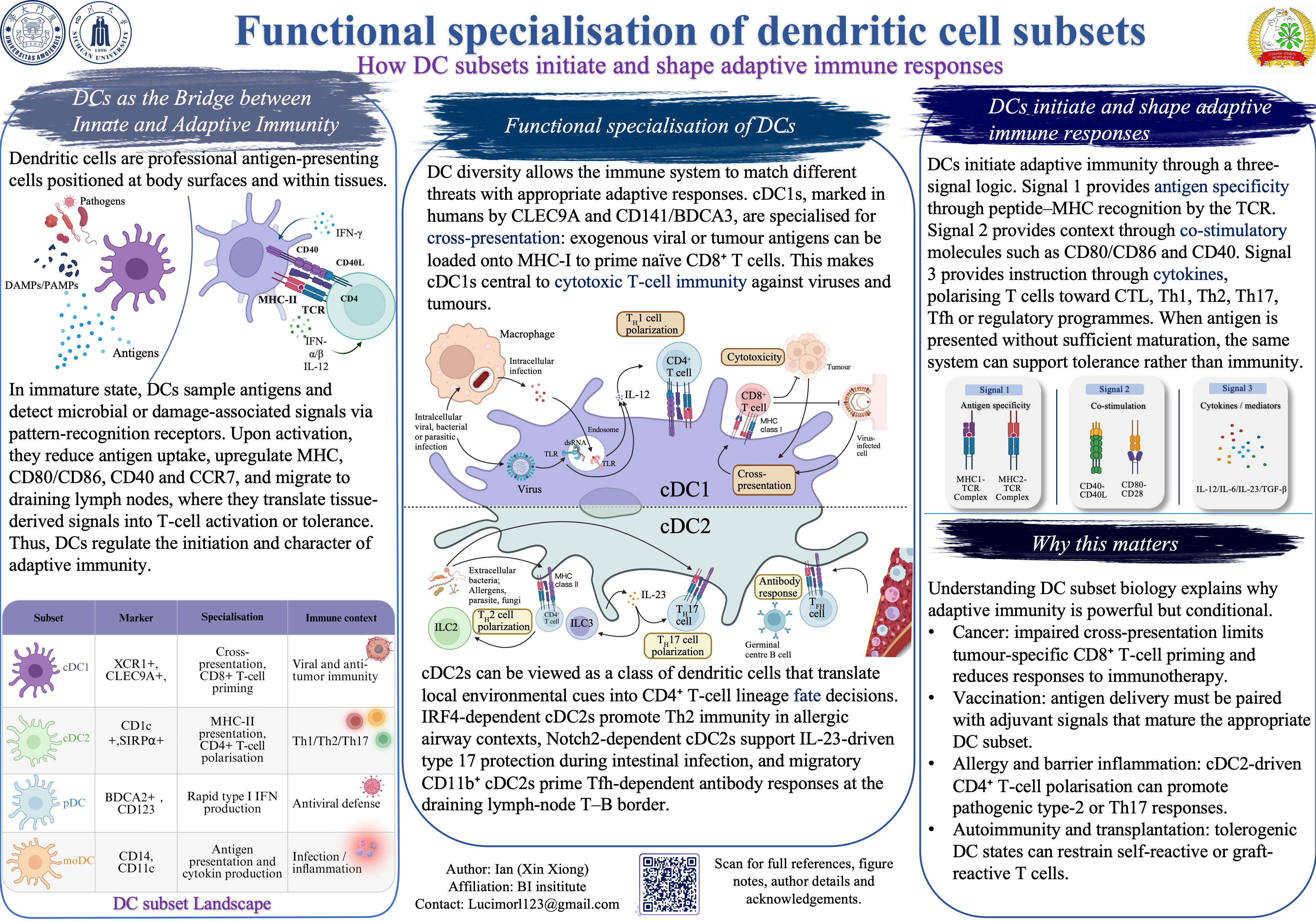
Full poster image for reference.
1. Poster question and core answer
Biological question
How do dendritic cell subsets convert tissue-derived danger signals into distinct adaptive immune responses?
One-sentence answer
Dendritic cells (DCs) do not simply display antigen: they integrate antigenic, inflammatory, spatial and maturation cues, then deliver Signal 1 through peptide–MHC, Signal 2 through co-stimulation, and Signal 3 through cytokines to determine whether T cells become cytotoxic, helper, follicular, regulatory or tolerant.
Take-home message
DC functional specialisation is best understood as a translation system:
Tissue cue → DC subset and maturation state → antigen-processing route → T-cell fate → immune outcome- cDC1 specialise in cross-presentation and CD8⁺ T-cell priming, supporting cytotoxic antiviral and anti-tumour immunity.
- cDC2 integrate local tissue cues and mainly instruct CD4⁺ T-cell fates, including Th2, Th17 and Tfh-associated responses.
- pDC rapidly produce type I interferons and condition antiviral immunity, but can also participate in immune regulation.
- moDC are best viewed as inflammation-induced APC-like states rather than a stable steady-state DC lineage equivalent to cDC1/cDC2.
- Tolerogenic DC states restrain inappropriate responses by promoting T-cell anergy, deletion or regulatory programmes.
2. Figure captions for the poster
Figure 1. DCs as innate–adaptive immune translators
Immature DCs are positioned at body surfaces and within tissues, where they continuously sample antigens and sense pathogen-associated or damage-associated signals through pattern-recognition receptors. Upon activation, they reduce antigen uptake, upregulate MHC molecules, CD80/CD86, CD40 and CCR7, then migrate to draining lymph nodes to initiate T-cell responses.
Figure 2. DC subset landscape
The table summarises major DC-related populations. cDC1 are associated with XCR1, CLEC9A and CD141/BDCA3 in humans and are specialised for cross-presentation. cDC2 are associated with CD1c and SIRPα and are more closely linked to MHC-II presentation and CD4⁺ T-cell polarisation. pDC are defined by rapid type I interferon production, whereas moDC usually arise in inflammatory contexts.
Figure 3. Functional specialisation of cDC1 and cDC2
The central mechanism diagram contrasts two major conventional DC programmes. cDC1 route exogenous or cell-associated antigens into MHC-I presentation and prime naïve CD8⁺ T cells, thereby supporting CTL-mediated antiviral and anti-tumour immunity. cDC2 use MHC-II presentation and tissue-derived inflammatory cues to shape CD4⁺ T-cell fates, including Th2, Th17 and Tfh-dependent antibody responses.
Figure 4. Three-signal model of DC-mediated T-cell priming
DCs initiate adaptive immunity through coordinated signals. Signal 1 provides antigen specificity through peptide–MHC recognition by the TCR. Signal 2 provides activation context through co-stimulatory interactions such as CD80/CD86–CD28 and CD40–CD40L. Signal 3 provides polarising instruction through cytokines such as IL-12, IL-6, IL-23, TGF-β and type I interferons.
Figure 5. Clinical and translational relevance
DC subset biology explains why adaptive immunity is powerful but conditional. Cancer immunity often requires effective cDC1-dependent cross-presentation and tumour-infiltrating DC–T-cell crosstalk. Vaccination requires antigen delivery plus appropriate adjuvant-driven DC maturation. Allergy, autoimmunity and transplantation show the opposite side of the same system: inappropriate or tolerogenic DC signals can determine whether adaptive immunity becomes pathogenic or restrained.
3. Image, diagram and resolution notes
- The poster diagrams are original schematic illustrations prepared for educational communication; they are conceptually based on the immunology literature rather than directly reproduced from any single published figure.
- The DC subset markers, functional annotations and signalling logic were checked against current review literature and course-level immunology concepts.
- All receptor labels, cytokine labels, arrows and figure captions should remain readable when displayed on a projector or when viewed as a full-size PDF.
- If the poster is printed, colours should be checked after PDF export because RGB screen colours may shift in CMYK printing.
4. Acknowledgements and tools
This poster was prepared for the BI Advanced Immunology poster and final presentation assessment. The scientific narrative was developed from course material, textbook-level immunology concepts and selected primary/review literature on dendritic cell ontogeny, antigen presentation, T-cell priming and immunotherapy.
Tools used: PowerPoint for layout assembly; BioRender-style vector illustration principles for scientific diagrams; AI-assisted tools for text refinement, layout planning and visual polishing. Final scientific content, terminology and figure logic were reviewed and edited by the author.
5. Concise research note
5.1 Why DCs are more than antigen-presenting cells
Dendritic cells are often introduced as professional antigen-presenting cells, but their biological role is broader. They determine not only whether adaptive immunity starts, but also what type of adaptive immunity is generated. This decision depends on where the antigen was captured, which PRRs were engaged, which DC subset processed the antigen, whether the DC matured, and which lymph-node microenvironment the DC entered.
The key logic is the three-signal model. Signal 1 selects antigen-specific T-cell clones through peptide–MHC recognition. Signal 2 determines whether the T cell interprets antigen recognition as activation rather than tolerance. Signal 3 instructs effector differentiation, such as CTL, Th1, Th2, Th17, Tfh or Treg programmes. Without sufficient maturation, antigen presentation can support tolerance instead of immunity.
5.2 cDC1: cross-presentation and cytotoxic immunity
cDC1 are the most clearly specialised conventional DC subset. In humans, they are commonly associated with XCR1, CLEC9A and CD141/BDCA3; in mice, they include lymphoid-resident CD8α⁺ DCs and migratory CD103⁺ DCs. Their development is strongly linked to the IRF8–BATF3 transcriptional programme.
The signature function of cDC1 is cross-presentation, in which exogenous or cell-associated antigens are loaded onto MHC-I and presented to CD8⁺ T cells. This is essential when viruses or tumours do not directly infect DCs but still need to be detected by cytotoxic T cells. cDC1 are therefore central to CTL priming, antiviral defence and anti-tumour immunity.
Mechanistically, cDC1 specialisation is not simply “better phagocytosis”. It involves specialised receptors and intracellular routing, including recognition of dead-cell material through CLEC9A/DNGR-1, regulation of phagosomal processing, antigen escape or transfer into MHC-I loading pathways, and efficient interaction with CD8⁺ T cells. In tumours, functional cDC1 can also provide re-stimulation and cytokine support that improve the quality of CD8⁺ T-cell responses.
5.3 cDC2: CD4⁺ T-cell polarisation and humoral immunity
cDC2 are more heterogeneous than cDC1. They are commonly associated with CD1c, SIRPα and FcεRIα in humans and with CD11b/SIRPα-associated phenotypes in mice. Their development and functional modules involve IRF4, with additional context-dependent regulation by factors such as Notch2 and KLF4.
The most concise way to describe cDC2 is that they translate tissue context into CD4⁺ T-cell fate decisions. In barrier tissues, cDC2 can promote type-2 immunity during allergic or helminth-associated contexts, type-17 immunity during mucosal bacterial or fungal challenge, and Tfh-associated programmes that support germinal-centre and antibody responses.
A key example for the poster is the role of migratory CD11b⁺ cDC2 in Tfh-dependent antibody responses. In mucosal immunisation models, migratory cDC2 transport antigen to the T–B border of draining lymph nodes, where they prime Tfh cells and thereby support downstream B-cell help and antibody production. This shows that DCs shape not only cellular immunity but also humoral immunity indirectly through Tfh induction.
5.4 pDC, moDC and tolerogenic DC states
pDC are best known for rapid type I interferon production after viral nucleic acid sensing, especially through TLR7 and TLR9. Their main contribution is therefore often upstream of full adaptive immunity: they establish an antiviral cytokine environment that conditions other DCs, NK cells and T cells. However, pDC can also participate in immune regulation, depending on activation state and tissue context.
moDC are frequently generated in vitro from monocytes and appear in inflamed tissues in vivo. They may present antigen and produce inflammatory cytokines, but they should not be treated as equivalent to steady-state cDC1 or cDC2. In a poster or viva answer, it is safer to describe moDC as inflammation-induced APC-like states that can amplify ongoing responses.
Tolerogenic DCs are also not a single fixed lineage. They are DC states with low co-stimulation and regulatory outputs such as IL-10, TGF-β, IDO or retinoic-acid-associated programmes. Their importance is that they reveal the other side of DC biology: the same antigen-presentation system that initiates immunity can also maintain self-tolerance and prevent immunopathology.
6. Comparison table
| DC subset/state | Representative markers or features | Main adaptive immune role | Poster emphasis |
|---|---|---|---|
| cDC1 | XCR1⁺, CLEC9A⁺, CD141/BDCA3⁺ in humans; BATF3/IRF8-dependent | Cross-presentation; naïve CD8⁺ T-cell priming; CTL and Th1-associated immunity | Viral and anti-tumour immunity |
| cDC2 | CD1c⁺, SIRPα⁺; IRF4/Notch2/KLF4-associated modules | MHC-II presentation; CD4⁺ T-cell polarisation; Th2/Th17/Tfh programmes | Tissue-context translation and antibody responses |
| pDC | CD123^hi^, BDCA2/CLEC4C⁺ in humans; TLR7/TLR9-rich | Rapid type I IFN production; antiviral conditioning; context-dependent regulation | Antiviral defence and immune regulation |
| moDC | Monocyte-derived; often CD14-associated in inflammatory contexts | Inflammatory antigen presentation and cytokine amplification | Infection/inflammation rather than steady-state priming |
| Tolerogenic DC state | Low co-stimulation; IL-10/TGF-β/IDO/RA-associated outputs | T-cell anergy, deletion or Treg induction | Autoimmunity, transplantation and peripheral tolerance |
7. Selected references
- Steinman RM, Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. Journal of Experimental Medicine. 1973;137(5):1142-1162. doi:10.1084/jem.137.5.1142.
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392(6673):245-252. doi:10.1038/32588.
- Guermonprez P, Valladeau J, Zitvogel L, Théry C, Amigorena S. Antigen Presentation and T Cell Stimulation by Dendritic Cells. Annual Review of Immunology. 2002;20:621-667. doi:10.1146/annurev.immunol.20.100301.064828.
- Trombetta ES, Mellman I. Cell Biology of Antigen Processing in Vitro and in Vivo. Annual Review of Immunology. 2005;23:975-1028. doi:10.1146/annurev.immunol.22.012703.104538.
- Hildner K, Edelson BT, Purtha WE, Diamond M, Matsushita H, et al. Batf3 deficiency reveals a critical role for CD8α⁺ dendritic cells in cytotoxic T cell immunity. Science. 2008;322(5904):1097-1100. doi:10.1126/science.1164206.
- Bachem A, Güttler S, Hartung E, Ebstein F, Schaefer M, et al. Expression of XCR1 characterizes the Batf3-dependent lineage of dendritic cells capable of antigen cross-presentation. Frontiers in Immunology. 2012;3:214. doi:10.3389/fimmu.2012.00214.
- Guilliams M, Ginhoux F, Jakubzick C, Naik SH, Onai N, et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nature Reviews Immunology. 2014;14(8):571-578. doi:10.1038/nri3712.
- Krishnaswamy JK, Gowthaman U, Zhang B, Mattsson J, Szeponik L, et al. Migratory CD11b⁺ conventional dendritic cells induce T follicular helper cell-dependent antibody responses. Science Immunology. 2017;2(18):eaam9169. doi:10.1126/sciimmunol.aam9169.
- Briseño CG, Satpathy AT, Davidson JT, Ferris ST, Durai V, et al. Notch2-dependent DC2s mediate splenic germinal center responses. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(42):10726-10731. doi:10.1073/pnas.1809925115.
- Reizis B. Plasmacytoid dendritic cells: development, regulation, and function. Annual Review of Immunology. 2019;37:163-183. doi:10.1146/annurev-immunol-041015-055531.
- Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic cells in cancer immunology and immunotherapy. Nature Reviews Immunology. 2020;20(1):7-24. doi:10.1038/s41577-019-0210-z. PMID:32908299.
- Cabeza-Cabrerizo M, Cardoso A, Minutti CM, Pereira da Costa M, Reis e Sousa C. Dendritic cells revisited. Annual Review of Immunology. 2021;39:131-166. doi:10.1146/annurev-immunol-061020-053707.
- Anderson DA III, Dutertre CA, Ginhoux F, Murphy KM. Genetic models of human and mouse dendritic cell development and function. Nature Reviews Immunology. 2021;21(2):101-115. doi:10.1038/s41577-020-00413-x.
- Owens T. T-cell costimulation: T cells themselves call the shots. Current Biology. 1996;6(1):32-35. doi:10.1016/S0960-9822(02)00415-3.
- Henry CM, Reis e Sousa C. DNGR-1-mediated cross-presentation of dead cell-associated antigens. Trends in Immunology. 2023;44(8):617-629. doi:10.1016/j.it.2023.05.006.
- Minutti CM, Piot C, Pereira da Costa M, Chakravarty P, Rogers N, et al. Distinct ontogenetic lineages dictate cDC2 heterogeneity. Nature Immunology. 2024;25(3):448-461. doi:10.1038/s41590-024-01745-9.